
Alle iLive-Inhalte werden medizinisch überprüft oder auf ihre Richtigkeit überprüft.
Wir haben strenge Beschaffungsrichtlinien und verlinken nur zu seriösen Medienseiten, akademischen Forschungseinrichtungen und, wenn möglich, medizinisch begutachteten Studien. Beachten Sie, dass die Zahlen in Klammern ([1], [2] usw.) anklickbare Links zu diesen Studien sind.
Wenn Sie der Meinung sind, dass einer unserer Inhalte ungenau, veraltet oder auf andere Weise bedenklich ist, wählen Sie ihn aus und drücken Sie Strg + Eingabe.
Hirnanhangsdrüse
Facharzt des Artikels

Zuletzt überprüft: 04.07.2025

Die Hypophyse (Hypophyse, S.glandula pituitaria) befindet sich in der Hypophysengrube der Sella turcica des Keilbeins und ist durch einen Fortsatz der Dura mater des Gehirns, der das Zwerchfell der Sella bildet, von der Schädelhöhle getrennt. Durch die Öffnung in diesem Zwerchfell ist die Hypophyse mit dem Infundibulum des Hypothalamus des Zwischenhirns verbunden. Die transversale Größe der Hypophyse beträgt 10-17 mm, die anteroposteriore 5-15 mm, die vertikale 5-10 mm. Die Masse der Hypophyse beträgt bei Männern etwa 0,5 g, bei Frauen 0,6 g. Die Hypophyse ist äußerlich von einer Kapsel umgeben.
Entsprechend der Entwicklung der Hypophyse aus zwei verschiedenen Rudimenten werden im Organ zwei Lappen unterschieden - der vordere und der hintere. Die Adenohypophyse oder der Vorderlappen (Adenohypophyse, s.lobus anterior) ist größer und macht 70-80% der Gesamtmasse der Hypophyse aus. Es ist dichter als der Hinterlappen. Im Vorderlappen unterscheidet man einen distalen Teil (Pars distalis), der den vorderen Teil der Hypophysengrube einnimmt, einen Zwischenteil (Pars intermedia), der sich an der Grenze zum Hinterlappen befindet, und einen knollenförmigen Teil (Pars tuberalis), der nach oben geht und mit dem Infundibulum des Hypothalamus verbunden ist. Aufgrund der Fülle an Blutgefäßen hat der Vorderlappen eine hellgelbe Farbe mit einem rötlichen Farbton. Das Parenchym des Hypophysenvorderlappens wird durch verschiedene Drüsenzellen repräsentiert, zwischen deren Strängen sich sinusförmige Blutkapillaren befinden. Die Hälfte (50 %) der Adenohypophysenzellen sind chromophile Adenozyten, deren Zytoplasma feinkörnige Granula aufweist, die sich gut mit Chromsalzen anfärben. Dies sind acidophile Adenozyten (40 % aller Adenohypophysenzellen) und basophile Adenozyten (10 %). Zu den basophilen Adenozyten zählen gonadotrope, kortikotrope und thyreotrope Endokrinozyten. Chromophobe Adenozyten sind klein, haben einen großen Kern und wenig Zytoplasma. Diese Zellen gelten als Vorläufer chromophiler Adenozyten. Die anderen 50 % der Adenohypophysenzellen sind chromophobe Adenozyten.

Die Neurohypophyse oder Hinterlappen (Neurohypophyse, s.lobus posterior) besteht aus dem Neurallappen (Lobus nervosus), der sich im hinteren Teil der Hypophysengrube befindet, und dem Trichter (Infundibulum), der sich hinter dem tuberösen Teil der Adenohypophyse befindet. Der Hinterlappen der Hypophyse wird von Neurogliazellen (Hypophysenzellen), Nervenfasern, die von den neurosekretorischen Kernen des Hypothalamus zur Neurohypophyse verlaufen, und neurosekretorischen Körperchen gebildet.
Die Hypophyse ist über Nervenfasern (Bahnen) und Blutgefäße funktionell mit dem Hypothalamus im Zwischenhirn verbunden, der die Aktivität der Hypophyse reguliert. Hypophyse und Hypothalamus werden zusammen mit ihren neuroendokrinen, vaskulären und nervlichen Verbindungen üblicherweise als Hypothalamus-Hypophysen-System bezeichnet.
Hormone der vorderen und hinteren Hypophyse beeinflussen viele Körperfunktionen, hauptsächlich über andere endokrine Drüsen. In der vorderen Hypophyse produzieren acidophile Adenozyten (Alpha-Zellen) das somatotrope Hormon (Wachstumshormon), das an der Regulierung von Wachstums- und Entwicklungsprozessen junger Organismen beteiligt ist. Kortikotrope Endokrinozyten sezernieren das adrenocorticotrope Hormon (ACTH), das die Sekretion von Steroidhormonen durch die Nebennieren stimuliert. Thyreotrope Endokrinozyten sezernieren das thyreotrope Hormon (TSH), das die Entwicklung der Schilddrüse beeinflusst und die Produktion ihrer Hormone aktiviert. Gonadotrope Hormone: follikelstimulierendes (FSH), luteinisierendes (LH) und Prolaktin – beeinflussen die sexuelle Reifung des Körpers, regulieren und stimulieren die Entwicklung der Follikel im Eierstock, den Eisprung, das Wachstum der Brustdrüsen und die Milchproduktion bei Frauen sowie den Prozess der Spermatogenese bei Männern. Diese Hormone werden von basophilen Adenozyten (Betazellen) produziert. Hier werden auch lipotrope Faktoren der Hypophyse ausgeschüttet, die die Mobilisierung und Verwertung von Fetten im Körper beeinflussen. Im Zwischenteil des Vorderlappens wird das Melanozyten-stimulierende Hormon gebildet, das die Bildung von Pigmenten – Melaninen – im Körper steuert.
Neurosekretorische Zellen der supraoptischen und paraventrikulären Kerne im Hypothalamus produzieren Vasopressin und Oxytocin. Diese Hormone werden über Axone, die den Hypothalamus-Hypophysen-Trakt bilden, zu den Zellen der Hypophyse transportiert. Von der Hypophyse gelangen diese Substanzen ins Blut. Das Hormon Vasopressin hat eine vasokonstriktive und antidiuretische Wirkung, weshalb es auch als antidiuretisches Hormon (ADH) bezeichnet wird. Oxytocin hat eine stimulierende Wirkung auf die Kontraktilität der Gebärmuttermuskulatur, erhöht die Milchsekretion der Milchdrüse, hemmt die Entwicklung und Funktion des Gelbkörpers und beeinflusst Tonusveränderungen der glatten (nicht quergestreiften) Muskulatur des Magen-Darm-Trakts.
Entwicklung der Hypophyse
Der Hypophysenvorderlappen entwickelt sich aus dem Epithel der dorsalen Wand der Mundhöhle in Form eines ringförmigen Auswuchses (Rathke-Tasche). Dieser ektodermale Vorsprung wächst zum Boden des zukünftigen dritten Ventrikels. Von der Unterseite der zweiten Hirnblase (dem zukünftigen Boden des dritten Ventrikels) wächst ein Prozess dorthin, aus dem sich der graue Tuberkel des Infundibulums und der Hypophysenhinterlappen entwickeln.
Gefäße und Nerven der Hypophyse
Von den inneren Halsschlagadern und Gefäßen des Großhirnarterienkreises werden die oberen und unteren Hypophysenarterien zur Hypophyse geleitet. Die oberen Hypophysenarterien verlaufen zum grauen Kern und zum Infundibulum des Hypothalamus, anastomosieren hier miteinander und bilden Kapillaren, die das Hirngewebe durchdringen – das primäre Hämokapillarnetz. Aus den langen und kurzen Schleifen dieses Netzes werden die Pfortadern gebildet, die zum Hypophysenvorderlappen gerichtet sind. Im Parenchym des Hypophysenvorderlappens teilen sich diese Venen in weite sinusförmige Kapillaren auf und bilden das sekundäre Hämokapillarnetz. Der Hypophysenhinterlappen wird hauptsächlich von der unteren Hypophysenarterie mit Blut versorgt. Zwischen den oberen und unteren Hypophysenarterien bestehen lange arterielle Anastomosen. Der Abfluss des venösen Blutes aus dem sekundären Hämokapillarnetz erfolgt über ein Venensystem, das in die Sinus cavernosus und intercavernosus der Dura mater des Gehirns mündet.
Sympathische Fasern, die zusammen mit Arterien in das Organ gelangen, sind an der Innervation der Hypophyse beteiligt. Postganglionäre sympathische Nervenfasern gehen vom Plexus der inneren Halsschlagader aus. Darüber hinaus befinden sich im Hypophysenhinterlappen zahlreiche Enden der Prozesse neurosekretorischer Zellen, die sich in den Kernen des Hypothalamus befinden.
Altersbedingte Merkmale der Hypophyse
Das durchschnittliche Gewicht der Hypophyse bei Neugeborenen beträgt 0,12 g. Das Gewicht des Organs verdoppelt sich im Alter von 10 Jahren und verdreifacht sich im Alter von 15 Jahren. Im Alter von 20 Jahren erreicht das Gewicht der Hypophyse sein Maximum (530-560 mg) und bleibt in den folgenden Altersperioden nahezu unverändert. Nach 60 Jahren ist eine leichte Gewichtsabnahme dieser endokrinen Drüse zu beobachten.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hypophysenhormone
Die Einheit der Nerven- und Hormonregulation im Körper wird durch die enge anatomische und funktionelle Verbindung von Hypophyse und Hypothalamus gewährleistet. Dieser Komplex bestimmt den Zustand und die Funktion des gesamten endokrinen Systems.
Die wichtigste endokrine Drüse, die eine Reihe von Peptidhormonen produziert, die die Funktion der peripheren Drüsen direkt regulieren, ist die Hypophyse. Es ist eine rötlich-graue, bohnenförmige Formation, die mit einer faserigen Kapsel mit einem Gewicht von 0,5 bis 0,6 g bedeckt ist. Das Gewicht variiert geringfügig je nach Geschlecht und Alter einer Person. Die allgemein anerkannte Unterteilung der Hypophyse in zwei Lappen, die sich in Entwicklung, Struktur und Funktion unterscheiden, bleibt bestehen: die vordere distale Adenohypophyse und die hintere Neurohypophyse. Der erste macht etwa 70% der Gesamtmasse der Drüse aus und wird üblicherweise in den distalen, infundibulären und intermediären Teil unterteilt, der zweite in den hinteren Teil oder Lappen und den Hypophysenstiel. Die Drüse befindet sich in der Hypophysengrube der Sella turcica des Keilbeins und ist über den Stiel mit dem Gehirn verbunden. Der obere Teil des Vorderlappens wird von der Sehnervenkreuzung und den Sehbahnen bedeckt. Die Blutversorgung der Hypophyse ist sehr reichlich und erfolgt über Äste der inneren Halsschlagader (die oberen und unteren Hypophysenarterien) sowie über Äste des Großhirnarterienkreises. Die oberen Hypophysenarterien sind an der Blutversorgung der Adenohypophyse beteiligt, die unteren an der Neurohypophyse und stehen mit den neurosekretorischen Enden der Axone der großzelligen Kerne des Hypothalamus in Kontakt. Erstere dringen in die Eminentia mediana des Hypothalamus ein, wo sie sich in ein Kapillarnetzwerk (den primären Kapillarplexus) aufteilen. Diese Kapillaren (mit denen die Axonenden der kleinen neurosekretorischen Zellen des mediobasalen Hypothalamus in Kontakt stehen) sammeln sich in Pfortadern, die entlang des Hypophysenstiels in das Parenchym der Adenohypophyse absteigen, wo sie sich erneut in ein Netzwerk sinusförmiger Kapillaren (den sekundären Kapillarplexus) aufteilen. So gelangt das Blut, das zuvor die Eminentia mediana des Hypothalamus passiert hat, wo es mit hypothalamischen adenohypophysotropen Hormonen (Releasing-Hormonen) angereichert wurde, in die Adenohypophyse.
Der Abfluss von mit Adenohypophysenhormonen gesättigtem Blut aus zahlreichen Kapillaren des sekundären Plexus erfolgt durch das Venensystem, das wiederum in die venösen Nebenhöhlen der Dura mater und dann in den allgemeinen Blutkreislauf mündet. Somit ist das Portalsystem der Hypophyse mit absteigender Blutflussrichtung vom Hypothalamus eine morphofunktionelle Komponente des komplexen Mechanismus der neurohumoralen Kontrolle der tropischen Funktionen der Adenohypophyse.
Die Hypophyse wird durch sympathische Fasern innerviert, die den Hypophysenarterien folgen. Sie entspringen postganglionären Fasern, die durch den inneren Karotisplexus verlaufen und mit den oberen Halsganglien verbunden sind. Eine direkte Innervation der Adenohypophyse durch den Hypothalamus erfolgt nicht. Der Hinterlappen erhält Nervenfasern aus den neurosekretorischen Kernen des Hypothalamus.
Die Adenohypophyse ist ein histologisch sehr komplexes Gebilde. Sie weist zwei Arten von Drüsenzellen auf – chromophobe und chromophile. Letztere werden wiederum in acidophile und basophile Zellen unterteilt (eine detaillierte histologische Beschreibung der Hypophyse findet sich im entsprechenden Abschnitt des Handbuchs). Es ist jedoch zu beachten, dass die von den Drüsenzellen, die das Parenchym der Adenohypophyse bilden, produzierten Hormone aufgrund ihrer Diversität in ihrer chemischen Natur teilweise unterschiedlich sind und die Feinstruktur der sezernierenden Zellen den Biosynthesemerkmalen jeder einzelnen entsprechen muss. Manchmal können jedoch in der Adenohypophyse Übergangsformen von Drüsenzellen beobachtet werden, die mehrere Hormone produzieren können. Es gibt Hinweise darauf, dass der Typ der Drüsenzellen der Adenohypophyse nicht immer genetisch bestimmt ist.
Unter dem Zwerchfell der Sella turcica befindet sich der trichterförmige Teil des Vorderlappens. Er umschließt den Hypophysenstiel und berührt den Tuberculum gravidum. Dieser Teil der Adenohypophyse ist durch das Vorhandensein von Epithelzellen und eine reichliche Blutversorgung gekennzeichnet. Er ist auch hormonell aktiv.
Der intermediäre (mittlere) Teil der Hypophyse besteht aus mehreren Schichten großer sekretorisch aktiver basophiler Zellen.
Die Hypophyse erfüllt über ihre Hormone verschiedene Funktionen. Ihr Vorderlappen produziert adrenocorticotrope (ACTH), schilddrüsenstimulierende (TSH), follikelstimulierende (FSH), luteinisierende (LH), lipotrope Hormone sowie das somatotrope Wachstumshormon (STO) und Prolaktin. Im Zwischenlappen wird das Melanozyten-stimulierende Hormon (MSH) synthetisiert, und im Hinterlappen reichern sich Vasopressin und Oxytocin an.
ACTH
Hypophysenhormone sind eine Gruppe von Protein- und Peptidhormonen sowie Glykoproteinen. Von den Hormonen des Hypophysenvorderlappens ist ACTH am besten untersucht. Es wird von basophilen Zellen produziert. Seine wichtigste physiologische Funktion ist die Stimulierung der Biosynthese und Sekretion von Steroidhormonen durch die Nebennierenrinde. ACTH zeigt außerdem melanozytenstimulierende und lipotrope Aktivität. 1953 wurde es in reiner Form isoliert. Später wurde seine chemische Struktur, bestehend aus 39 Aminosäureresten, beim Menschen und einer Reihe von Säugetieren entschlüsselt. ACTH ist nicht speziesspezifisch. Bislang wurden sowohl das Hormon selbst als auch verschiedene Fragmente seines Moleküls, die aktiver als natürliche Hormone sind, chemisch synthetisiert. Die Struktur des Hormons besteht aus zwei Abschnitten der Peptidkette, von denen einer die Erkennung und Bindung von ACTH an den Rezeptor gewährleistet, während der andere für eine biologische Wirkung sorgt. Die Bindung an den ACTH-Rezeptor erfolgt offenbar durch die Wechselwirkung der elektrischen Ladungen des Hormons und des Rezeptors. Die Rolle des biologischen Effektors von ACTH übernimmt das Fragment des 4-10-Moleküls (Met-Glu-His-Phen-Arg-Tri-Tri).
Die melanozytenstimulierende Wirkung von ACTH beruht auf der N-terminalen Region im Molekül, die aus 13 Aminosäureresten besteht und die Struktur des alpha-melanozytenstimulierenden Hormons wiederholt. Diese Region enthält auch ein Heptapeptid, das in anderen Hypophysenhormonen vorkommt und adrenocorticotrope, melanozytenstimulierende und lipotrope Wirkungen besitzt.
Als Schlüsselmoment der ACTH-Wirkung gilt die Aktivierung des Enzyms Proteinkinase im Zytoplasma unter Beteiligung von cAMP. Phosphorylierte Proteinkinase aktiviert das Enzym Esterase, das Cholesterinester in Fetttröpfchen in eine freie Substanz umwandelt. Das im Zytoplasma durch Phosphorylierung von Ribosomen synthetisierte Protein stimuliert die Bindung von freiem Cholesterin an Cytochrom P-450 und dessen Transfer von Lipidtröpfchen zu Mitochondrien, wo alle Enzyme vorhanden sind, die die Umwandlung von Cholesterin in Kortikosteroide gewährleisten.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Schilddrüsenstimulierendes Hormon
TSH – Thyreotropin – der Hauptregulator der Entwicklung und Funktion der Schilddrüse, der Prozesse der Synthese und Sekretion von Schilddrüsenhormonen. Dieses komplexe Protein – Glykoprotein – besteht aus Alpha- und Beta-Untereinheiten. Die Struktur der ersten Untereinheit stimmt mit der Alpha-Untereinheit des luteinisierenden Hormons überein. Darüber hinaus stimmt sie bei verschiedenen Tierspezies weitgehend überein. Die Sequenz der Aminosäurereste in der Beta-Untereinheit des menschlichen TSH wurde entschlüsselt und besteht aus 119 Aminosäureresten. Es ist festzustellen, dass die Beta-Untereinheiten von menschlichem und Rinder-TSH in vielerlei Hinsicht ähnlich sind. Die biologischen Eigenschaften und die Art der biologischen Aktivität von Glykoproteinhormonen werden durch die Beta-Untereinheit bestimmt. Sie gewährleistet auch die Interaktion des Hormons mit Rezeptoren in verschiedenen Zielorganen. Allerdings zeigt die Beta-Untereinheit bei den meisten Tieren erst dann spezifische Aktivität, wenn sie sich mit der Alpha-Untereinheit verbindet, die als eine Art Aktivator des Hormons fungiert. Letztere induziert mit gleicher Wahrscheinlichkeit luteinisierende, follikelstimulierende und thyreotrope Aktivitäten, die durch die Eigenschaften der Beta-Untereinheit bestimmt werden. Die festgestellte Ähnlichkeit lässt den Schluss zu, dass diese Hormone im Laufe der Evolution aus einem gemeinsamen Vorgänger entstanden sind. Die Beta-Untereinheit bestimmt auch deren immunologische Eigenschaften. Es wird angenommen, dass die Alpha-Untereinheit die Beta-Untereinheit vor der Wirkung proteolytischer Enzyme schützt und ihren Transport von der Hypophyse zu den peripheren Zielorganen erleichtert.
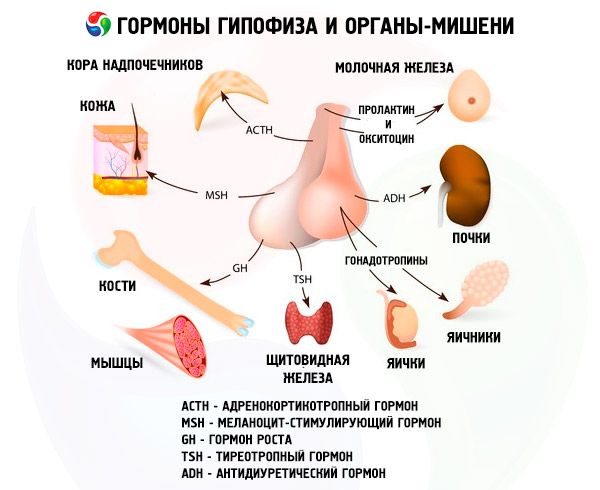
Gonadotrope Hormone
Gonadotropine kommen im Körper als LH und FSH vor. Der funktionelle Zweck dieser Hormone beschränkt sich im Allgemeinen auf die Gewährleistung von Fortpflanzungsprozessen bei Personen beiderlei Geschlechts. Sie sind wie TSH komplexe Proteine – Glykoproteine. FSH induziert die Reifung der Follikel in den Eierstöcken von Frauen und stimuliert die Spermatogenese bei Männern. LH verursacht bei Frauen einen Follikelbruch mit der Bildung des Gelbkörpers und stimuliert die Sekretion von Östrogenen und Progesteron. Bei Männern beschleunigt dasselbe Hormon die Entwicklung des interstitiellen Gewebes und die Sekretion von Androgenen. Die Wirkungen der Gonadotropine sind voneinander abhängig und treten synchron auf.
Die Dynamik der Gonadotropinsekretion bei Frauen verändert sich während des Menstruationszyklus und wurde ausreichend detailliert untersucht. In der präovulatorischen (follikulären) Phase des Zyklus ist der LH-Gehalt relativ niedrig, während der FSH-Spiegel erhöht ist. Mit zunehmender Reifung des Follikels nimmt die Östradiolsekretion zu, was eine erhöhte Gonadotropinproduktion durch die Hypophyse und das Auftreten sowohl von LH- als auch von FSH-Zyklen fördert, d. h. Sexualsteroide stimulieren die Gonadotropinsekretion.
Die Struktur von LH ist inzwischen geklärt. Wie TSH besteht es aus zwei Untereinheiten: a und b. Die Struktur der Alpha-Untereinheit von LH ist bei verschiedenen Tierarten weitgehend identisch und entspricht der Struktur der Alpha-Untereinheit von TSH.
Die Struktur der Beta-Untereinheit von LH unterscheidet sich deutlich von der Struktur der Beta-Untereinheit von TSH, obwohl sie vier identische Abschnitte der Peptidkette mit jeweils 4–5 Aminosäureresten aufweist. In TSH befinden sie sich an den Positionen 27–31, 51–54, 65–68 und 78–83. Da die Beta-Untereinheit von LH und TSH die spezifische biologische Aktivität der Hormone bestimmt, kann angenommen werden, dass homologe Abschnitte in der Struktur von LH und TSH die Verbindung der Beta-Untereinheiten mit der Alpha-Untereinheit sicherstellen und Abschnitte unterschiedlicher Struktur für die Spezifität der biologischen Aktivität der Hormone verantwortlich sein sollten.
Natives LH ist gegenüber der Wirkung proteolytischer Enzyme sehr beständig, die Beta-Untereinheit wird jedoch rasch durch Chymotrypsin gespalten, und die A-Untereinheit kann durch das Enzym nur schwer hydrolysiert werden, d. h. sie spielt eine schützende Rolle, indem sie Chymotrypsin den Zugriff auf die Peptidbindungen verwehrt.
Zur chemischen Struktur von FSH liegen noch keine eindeutigen Erkenntnisse vor. Wie LH besteht FSH aus zwei Untereinheiten, wobei sich die Beta-Untereinheit von FSH von der von LH unterscheidet.
Prolaktin
Ein weiteres Hormon, Prolaktin (laktogenes Hormon), spielt eine aktive Rolle bei Reproduktionsprozessen. Die wichtigsten physiologischen Eigenschaften von Prolaktin bei Säugetieren äußern sich in der Stimulation der Entwicklung der Brustdrüsen und der Milchproduktion, des Wachstums der Talgdrüsen und der inneren Organe. Es fördert die Wirkung von Steroiden auf die sekundären Geschlechtsmerkmale bei Männern, stimuliert die sekretorische Aktivität des Gelbkörpers bei Mäusen und Ratten und ist an der Regulierung des Fettstoffwechsels beteiligt. Prolaktin hat in den letzten Jahren als Regulator des mütterlichen Verhaltens große Aufmerksamkeit erhalten; diese Polyfunktionalität erklärt sich durch seine evolutionäre Entwicklung. Es ist eines der ältesten Hypophysenhormone und kommt sogar bei Amphibien vor. Inzwischen ist die Struktur von Prolaktin bei einigen Säugetierarten vollständig entschlüsselt. Bis vor kurzem äußerten Wissenschaftler jedoch Zweifel an der Existenz eines solchen Hormons beim Menschen. Viele glaubten, dass seine Funktion vom Wachstumshormon übernommen wird. Nun liegen überzeugende Beweise für das Vorhandensein von Prolaktin beim Menschen vor und seine Struktur wurde teilweise entschlüsselt. Prolaktinrezeptoren binden aktiv Wachstumshormon und Plazentalaktogen, was auf einen einheitlichen Wirkmechanismus der drei Hormone hindeutet.
Somatotropin
Das Wachstumshormon Somatotropin hat ein noch breiteres Wirkungsspektrum als Prolaktin. Wie Prolaktin wird es von azidophilen Zellen der Adenohypophyse produziert. STH stimuliert das Skelettwachstum, aktiviert die Proteinbiosynthese, wirkt fettmobilisierend und fördert die Körpergrößenzunahme. Darüber hinaus koordiniert es Stoffwechselprozesse.
Die Beteiligung des Hormons an letzterer wird durch die Tatsache bestätigt, dass seine Sekretion durch die Hypophyse beispielsweise bei einem Abfall des Blutzuckerspiegels stark ansteigt.
Die chemische Struktur dieses menschlichen Hormons ist inzwischen vollständig geklärt – sie besteht aus 191 Aminosäuren. Seine Primärstruktur ähnelt der des Chorion-Somatomammotropins oder des Plazentalaktogens. Diese Daten deuten auf eine signifikante evolutionäre Ähnlichkeit der beiden Hormone hin, obwohl sie Unterschiede in der biologischen Aktivität aufweisen.
Es muss die hohe Speziesspezifität des betreffenden Hormons betont werden – beispielsweise ist STH tierischen Ursprungs beim Menschen inaktiv. Dies erklärt sich sowohl durch die Reaktion zwischen den STH-Rezeptoren von Mensch und Tier als auch durch die Struktur des Hormons selbst. Derzeit werden Studien durchgeführt, um aktive Zentren mit biologischer Aktivität in der komplexen Struktur von STH zu identifizieren. Dabei werden auch einzelne Fragmente des Moleküls mit anderen Eigenschaften untersucht. So wurde beispielsweise nach der Hydrolyse von menschlichem STH durch Pepsin ein Peptid isoliert, das aus 14 Aminosäureresten besteht und dem Molekülabschnitt 31–44 entspricht. Es hatte keine wachstumsfördernde Wirkung, übertraf das native Hormon jedoch erheblich in seiner lipotropen Aktivität. Im Gegensatz zu ähnlichen tierischen Hormonen weist das menschliche Wachstumshormon eine signifikante laktogene Aktivität auf.
Die Adenohypophyse synthetisiert viele Peptid- und Proteinsubstanzen mit fettmobilisierender Wirkung, und die hypophysären Tropenhormone – ACTH, STH, TSH und andere – wirken lipotrop. In den letzten Jahren wurde insbesondere auf beta- und y-lipotrope Hormone (LPG) geachtet. Die biologischen Eigenschaften von Beta-LPG sind am besten untersucht; neben der lipotropen Aktivität hat es auch eine melanozyt-stimulierende, corticotropin-stimulierende und hypokalzämische Wirkung und erzeugt zudem eine insulinähnliche Wirkung.
Die Primärstruktur von Schaf-LPG (90 Aminosäurereste), lipotropen Hormonen von Schweinen und Rindern, ist inzwischen entschlüsselt. Dieses Hormon ist artspezifisch, obwohl die Struktur des zentralen Bereichs von Beta-LPG bei verschiedenen Arten gleich ist. Sie bestimmt die biologischen Eigenschaften des Hormons. Ein Fragment dieses Bereichs findet sich in der Struktur von Alpha-MSH, Beta-MSH, ACTH und Beta-LPG. Es wird vermutet, dass diese Hormone im Laufe der Evolution aus demselben Vorläufer entstanden sind. γ-LPG hat eine schwächere lipotrope Aktivität als Beta-LPG.
Melanozyten-stimulierendes Hormon
Dieses im Zwischenlappen der Hypophyse synthetisierte Hormon stimuliert in seiner biologischen Funktion die Biosynthese des Hautpigments Melanin und fördert eine Zunahme der Größe und Anzahl der Pigmentzellen Melanozyten in der Haut von Amphibien. Diese Eigenschaften von MSH werden bei biologischen Tests des Hormons genutzt. Es gibt zwei Arten des Hormons: Alpha- und Beta-MSH. Es wurde gezeigt, dass Alpha-MSH keine Spezies-Spezifität aufweist und bei allen Säugetieren die gleiche chemische Struktur hat. Sein Molekül ist eine Peptidkette, die aus 13 Aminosäureresten besteht. Beta-MSH hingegen ist Spezies-Spezifität und seine Struktur unterscheidet sich bei verschiedenen Tieren. Bei den meisten Säugetieren besteht das Beta-MSH-Molekül aus 18 Aminosäureresten und nur beim Menschen ist es am Aminosäureende um vier Aminosäurereste verlängert. Es ist zu beachten, dass Alpha-MSH eine gewisse adrenocorticotrope Aktivität aufweist und seine Wirkung auf das Verhalten von Tieren und Menschen inzwischen nachgewiesen wurde.
Oxytocin und Vasopressin
Vasopressin und Oxytocin, die im Hypothalamus synthetisiert werden, reichern sich im Hypophysenhinterlappen an: Vasopressin in den Neuronen des Nucleus supraopticus und Oxytocin im Nucleus paraventriculatorius. Anschließend werden sie in die Hypophyse transportiert. Es ist hervorzuheben, dass die Vorstufe des Hormons Vasopressin zuerst im Hypothalamus synthetisiert wird. Gleichzeitig werden dort die Proteine Neurophysin Typ 1 und 2 produziert. Ersteres bindet Oxytocin, letzteres Vasopressin. Diese Komplexe wandern in Form neurosekretorischer Granula im Zytoplasma entlang des Axons und erreichen den Hypophysenhinterlappen, wo die Nervenfasern in der Gefäßwand enden und der Inhalt der Granula ins Blut gelangt. Vasopressin und Oxytocin sind die ersten Hypophysenhormone mit vollständig etablierter Aminosäuresequenz. In ihrer chemischen Struktur sind sie Nonapeptide mit einer Disulfidbrücke.
Die betrachteten Hormone erzeugen verschiedene biologische Wirkungen: Sie stimulieren den Transport von Wasser und Salzen durch Membranen, wirken vasopressorisch, verstärken die Kontraktionen der glatten Gebärmuttermuskulatur während der Geburt und erhöhen die Sekretion der Brustdrüsen. Es ist zu beachten, dass Vasopressin eine höhere antidiuretische Wirkung als Oxytocin hat, während letzteres eine stärkere Wirkung auf Gebärmutter und Brustdrüse hat. Der Hauptregulator der Vasopressinsekretion ist der Wasserverbrauch; in den Nierentubuli bindet es an Rezeptoren in den Zytoplasmamembranen und aktiviert anschließend das Enzym Adenylatcyclase in diesen. Verschiedene Teile des Moleküls sind für die Bindung des Hormons an den Rezeptor und für die biologische Wirkung verantwortlich.
Die Hypophyse, die über den Hypothalamus mit dem gesamten Nervensystem verbunden ist, vereint das endokrine System zu einem funktionellen Ganzen und trägt zur Konstanz des inneren Körpers (Homöostase) bei. Innerhalb des endokrinen Systems erfolgt die homöostatische Regulation nach dem Prinzip der Rückkopplung zwischen dem Hypophysenvorderlappen und den Zieldrüsen (Schilddrüse, Nebennierenrinde, Keimdrüsen). Ein Überschuss des von der Zieldrüse produzierten Hormons hemmt, ein Mangel stimuliert die Sekretion und Freisetzung des entsprechenden Tropenhormons. Der Hypothalamus ist in das Rückkopplungssystem eingebunden. Darin befinden sich die Rezeptorzonen, die für die Hormone der Zieldrüsen empfindlich sind. Durch die spezifische Bindung an im Blut zirkulierende Hormone und die Veränderung der Reaktion in Abhängigkeit von der Hormonkonzentration übertragen hypothalamische Rezeptoren ihre Wirkung auf die entsprechenden hypothalamischen Zentren, die die Arbeit der Adenohypophyse koordinieren und hypothalamische adenohypophysiotrope Hormone freisetzen. Daher sollte der Hypothalamus als neuroendokrines Gehirn betrachtet werden.
Использованная литература